It is a profound realization that the entire history, biological potential, and physical blueprint of a human being are stored within a single type of molecule found in nearly every cell of our bodies. This molecule—Deoxyribonucleic acid, or DNA—was first identified as “Nuclein” by Friedrich Miescher in 1869 after he isolated it from the nuclei of pus cells. While the term “nucleic acid” was later coined by Richard Altman, it was the scientist Zacharias who eventually gave this genetic archive its modern name: DNA.
While James Watson and Francis Crick, building on the X-ray crystallography of Maurice Wilkins and Rosalind Franklin, famously proposed the double helix model in 1953, the molecule is far more than just a spiral ladder. It is a chemical masterpiece of stability and precision, a living archive that defines the boundary between a bacterium and a human.
1. It’s Not Just a Shape—It’s an Electric Molecule
While we often focus on the iconic geometry of the helix, the molecule’s “electric personality” is what truly governs its life-sustaining functions. DNA architectures itself as a fundamentally acidic substance with a persistent negative charge. This is not a mere chemical byproduct; it is a critical feature of its stability.
This negative charge arises from the phosphate groups that form the backbone of each polynucleotide chain. Each phosphate group forms a covalent phosphoester bond with the 5′ carbon of the deoxyribose sugar, while the opposite end of the chain is marked by a 3′ hydroxyl group. By releasing protons, these phosphate groups ensure the molecule remains negatively charged, allowing it to interact precisely with proteins in the nucleus. Furthermore, DNA possesses the “hidden” physical property of absorbing ultraviolet rays at a specific wavelength of 2600Å, a signature used by researchers to quantify the very essence of life in a lab.
“DNA as an acidic substance, present in nucleus, was first identified by Friedrich Miescher in 1869.“
2. The Perfect Geometry of the “Uneven” Pair
The “hallmark” of the double helix is its complementary base pairing, a system of mathematical and spatial perfection. The nitrogenous bases are divided into two classes: the two-ringed Purines (Adenine and Guanine) and the single-ringed Pyrimidines (Cytosine and Thymine).
Nature ensures a uniform distance of approximately 20Å between the two strands by adhering to a strict rule: a purine must always pair with a pyrimidine. This creates a “rung” that is consistently three rings wide (2 from the purine and 1 from the pyrimidine), preventing the helix from bulging or narrowing.
| Base Pair | Type | Hydrogen Bonds |
|---|---|---|
| Adenine (A) – Thymine (T) | Purine–Pyrimidine | 2 |
| Guanine (G) – Cytosine (C) | Purine–Pyrimidine | 3 |
This molecular dance is secured by specific hydrogen bonds:
- Adenine (A) binds to Thymine (T) via two hydrogen bonds.
- Guanine (G) binds to Cytosine (C) via three hydrogen bonds.
Beyond these bonds, the structure is stabilized by the way the base pairs stack one over the other like plates, a physical arrangement that confers immense structural integrity. Even the chemical attachments are precise: the nitrogenous bases connect to the sugar via an N-glycosidic linkage, specifically at the N1 position for pyrimidines and the N9 position for purines. In a brilliant stroke of chemical nuance, nature distinguishes DNA from its cousin RNA by using Thymine (chemically known as 5-methyl uracil), a modification that adds a layer of stability to our permanent genetic records.
3. The Species-Specific “Base Code”
While the nitrogenous alphabet (A, T, G, C) is universal, the “math” of their proportions defines the species. This is governed by Chargaff’s Equivalency Rule, which states that in double-stranded DNA, the amount of purines equals the amount of pyrimidines ([A] + [G] = [T] + [C]).
| Organism Type | Base Composition | Base Ratio |
|---|---|---|
| Eukaryotes | A+T rich | >1 |
| Prokaryotes | G+C rich | <1 |
However, the Base Ratio—the ratio of (A + T) / (G + C)—is a constant that acts as a biological fingerprint:
- Eukaryotes (including humans): Possess “A-T type” DNA, where the A+T content exceeds G+C, resulting in a base ratio greater than one.
- Prokaryotes (bacteria): Often possess “G-C type” DNA, where the G+C content is higher, resulting in a base ratio less than one.
4. DNA’s “Thermostat”: The Unzipping Act
DNA is remarkably resilient, capable of “breaking” its structure to allow the code to be read and then snapping back together with perfect fidelity. This is known as Denaturation and Renaturation.
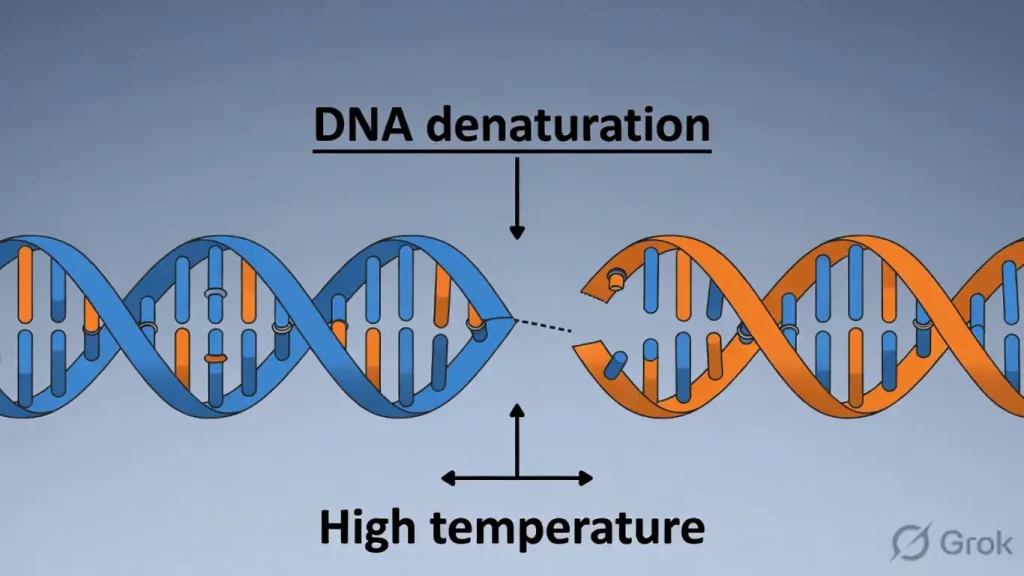
When exposed to high temperatures (92∘C−94∘C), the hydrogen bonds between the strands break, causing the helix to unzip. Because Guanine and Cytosine are held together by three hydrogen bonds rather than two, the Melting Point (Tm) of a DNA molecule is directly proportional to its G-C content. Consequently, prokaryotic DNA usually has a higher T_m than eukaryotic DNA. When the environment cools, the strands naturally find their complements and recoil into their original shape—a testament to the molecule’s inherent “memory” for its own geometry.
5. From Viruses to Humans: The Scale of Complexity
The volume of information packed into these microscopic strands scales dramatically as life moves from simple invaders to complex organisms. The jump in data is staggering:
- ϕ× 174 bacteriophage: 5,386 nucleotides (a unique virus utilizing single-stranded DNA).
- λ bacteriophage: 48,502 base pairs.
- E. coli: 4.6×10^6 base pairs.
- Human Haploid cell: 3.3×10^9 base pairs.
- Human Diploid cell: 6.6 × 10^9 base pairs.
To move from the 5,386 instructions of a virus to the 6.6 billion base pairs in a single human cell is to witness the incredible expansion of biological data required to weave the fabric of human consciousness and complexity.
6. Conclusion: The Living Archive
The structural brilliance of DNA—from its negatively charged phosphate backbone to the mathematical precision of its 3-ringed rungs—reveals a molecule perfectly tuned for its role as a living archive. It is a structure that is both fragile enough to be “unzipped” for the machinery of life and resilient enough to preserve its message across millennia.
Every cell in your body currently carries 6.6 billion base pairs of “instructional” history. As we peer deeper into the stacking of these bases and the electrical charges that hold them, we must ask: what other secrets remain hidden in the geometry of the helix? The future of biology lies in finally learning to read every page of the archive we carry in our pockets—and our nuclei.
Follow us: EcoGeneZap

